Глаз
Хруста́лик
(лат. lens) — прозрачное тело, расположенное внутри глазного яблока напротив зрачка; является биологической линзой, хрусталик составляет важную часть светопреломляющего аппарата глаза.
Хрусталик представляет собой прозрачное двояковыпуклое округлое эластичное образование, циркулярно фиксированное к цилиарному телу. Задняя поверхность хрусталика прилегает к стекловидному телу, спереди от него находятся радужка и передняя и задняя камеры.
Размеры и оптические свойства
Максимальная толщина хрусталика взрослого человека примерно 3,6—5 мм (в зависимости от напряжения аккомодации), его диаметр около 9—10 мм. Радиус кривизны передней поверхности хрусталика в покое аккомодации равен 10 мм, а задней — 6 мм, при максимальном напряжении аккомодации передний и задний радиус сравниваются, уменьшаясь до 5,33 мм.
Показатель преломления хрусталика неоднороден по толщине и в среднем составляет 1,386 или 1,406 (ядро) также в зависимости от состояния аккомодации.
В покое аккомодации преломляющая сила хрусталика составляет в среднем 19,11 диоптрий, при максимальном напряжении аккомодации — 33,06 дптр.
У новорождённых хрусталик почти шаровидный, имеет мягкую консистенцию и преломляющую силу до 35,0 дптр. Дальнейший рост его происходит, в основном, за счёт увеличения диаметра.
Симптоматика заболеваний хрусталика
С возрастом структура хрусталика меняется: он остановится более плотным и хуже реагирует на натяжение связочного аппарата. Из-за этого у Пациентов старше 40 лет нередко появляются жалобы на снижение зрения вблизи, т. е. развивается пресбиопия.
Возрастные изменения, нарушение обмена веществ приводят к потере прозрачности хрусталика — образуется катаракта. Самый частый симптом этого заболевания — помутнение в глазу: изображение становится желтоватым и тусклым. Возникает ощущение, что все вокруг видно через целлофановую пленку. Могут появиться ореолы при взгляде на источники света.
Возрастные помутнения хрусталика прогрессируют довольно медленно, вплоть до нескольких десятков лет. Иногда причиной катаракты является не возраст, а длительные воспаление глаз или глаукома (повышение внутриглазного давления). Спровоцировать помутнение хрусталика могут и травмы глаза.
Не всегда заболевания дают знать о себе яркими симптомами, поэтому мы советуем при любых, даже самых незначительных изменениях обратиться к опытному врачу-офтальмологу.
Гистологическое строение
В хрусталике выделяют капсулу
(сумку),
капсулярный эпителий
и
основное вещество хрусталика
.
Капсула
Снаружи хрусталик покрыт тонкой эластичной бесструктурной капсулой, которая представляет собой однородную прозрачную оболочку, сильно преломляющую свет и защищающую хрусталик от воздействия различных патологических факторов. Капсула при помощи ресничного пояска прикрепляется к ресничному телу.
Толщина капсулы хрусталика по всей его поверхности неодинакова: спереди часть капсулы толще, чем сзади (соответственно 0,008—0,02 и 0,002—0,004 мм), это обусловлено тем, что на передней поверхности под капсулой располагается одиночный слой эпителиальных клеток.
Наибольшей толщины капсула достигает в двух концентричных экватору её поясах — переднем (находится в 1 мм кнутри от места прикрепления передних волокон ресничного пояска) и заднем (кнутри от места заднего прикрепления ресничного пояска). Наименьшая толщина капсулы — в области заднего полюса хрусталика.
Эпителий
Эпителий хрусталика характеризуется как однослойный плоский неороговевающий; главными его функциями являются трофическая, камбиальная и барьерная.
Эпителиальные клетки, соответствующие центральной зоне капсулы (напротив зрачка), уплощены и плотно прилегают друг к другу. Здесь практически не происходит деление клеток.
По мере продвижения от центра к периферии наблюдается уменьшение размера эпителиальных клеток, усиление их митотической активности, а также относительное увеличение высоты клеток так, что в области экватора эпителий хрусталика практически превращается в призматический, образуя ростковую зону хрусталика. Здесь происходит образование так называемых волокон хрусталика.
Вещество хрусталика
Основная масса хрусталика образована волокнами, которые представляют собой клетки эпителия, вытянутые в длину. Каждое волокно представляет собой прозрачную шестиугольную призму. Вещество хрусталика, образованное белком кристаллином, совершенно прозрачно и так же, как другие компоненты светопреломляющего аппарата, лишено сосудов и нервов. Центральная, более плотная часть хрусталика, утратила ядро, укоротилась, и при наложении на другое волокно стала называться ядром
, в то время, как периферическая часть образует менее плотную
кору
.
В процессе внутриутробного развития хрусталик получает питание от стекловидной артерии. Во взрослом состоянии питание хрусталика всецело зависит от стекловидного тела и водянистой влаги.
Диагностика и лечение заболеваний хрусталика глаза
Для оценки состояния и работы хрусталика используют метод биомикроскопии (бесконтактного обследования с помощью щелевой лампы). Так врач-офтальмолог определяет размер хрусталика, степень его прозрачности, выявляет наличие и расположение помутнений.
Для лечения катаракты врачи Глазной клиники доктора Беликовой используют самый современный, безопасный и быстрый на сегодняшний день метод — ультразвуковую факоэмульсификацию. Заменив помутневший хрусталик на искусственную ИОЛ (интраокулярную линзу), можно восстановить зрение до 100% и избавиться от очков навсегда!
Специалистам
БИОМЕХАНИКА АККОМОДАЦИИ
Глава из книги «Биомеханика глаза»
(Е.Н.Иомдина, С.М.Бауэр, К.Е.Котляр)
Представлены результаты российских и зарубежных исследований, посвященных изучению биомеханических структур глаза. Показаны возможности использования результатов исследований как для понимания патогенеза различных заболеваний глаз, так и в клинической практике.
ПОЛНЫЙ ТЕКСТ КНИГИ НА https://sabar.eye-portal.ru
Биомеханика глаза — не только одно из научных направлений офтальмологии, позволяющих лучше понять работу глаза в норме и патологии, но также эффективный инструмент для разработки новых средств диагностики, оптической и хирургической коррекции зрительных нарушений . В данной главе представлены основные клинические приложения биомеханических исследований различных структур и систем органа зрения.
Биомеханика аккомодации
Аккомодация – основа нормального зрительного восприятия – представляет собой биомеханическую систему (с обратной связью) оптической настройки глаза для четкого видения предметов, находящихся на различных расстояниях, которая включает, прежде всего, хрусталик, его связочный аппарат (зонулярные волокна), цилиарную мышцу и хориоидею. В биомеханизм аккомодации в той или иной степени вовлечены и другие структуры глазного яблока, а именно, радужка (сфинктер зрачка), глазодвигательные мышцы, стекловидное тело, роговица, веки (руководство по аккомодации).
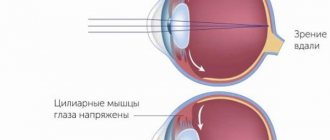
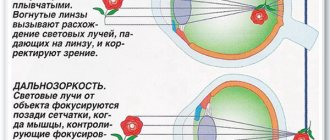
На сегодняшний день основополагающей теорией, описывающей механизм аккомодации, является гипотеза Германа фон Гельмгольца, согласно которой для получения четкого изображения расположенных на близком расстоянии предметов в человеческом глазу происходят следующие изменения: сокращается цилиарная мышца, происходит сужение зрачка, уменьшается глубина передней камеры, хрусталик смещается несколько кпереди и книзу, ослабевает натяжение цинновых связок, уменьшается радиус кривизны передней и задней (в меньшей степени) поверхностей хрусталика, что приводит к увеличению его преломляющей силы и усилению динамической рефракции глаза (Helmholtz H.v., 1855).
С момента опубликования точки зрения Гельмгольца на механизм аккомодации не прекращаются попытки оспорить, видоизменить или дополнить его теорию, поскольку не все клинические ситуации могут быть описаны с позиции этой теории.
Современные теории аккомодации можно разделить на интракапсулярные и экстракапсулярные. Первую группу теорий объединяет общий биомеханический принцип: при ослаблении связочного аппарата (зонулярных волокон) упругие силы капсулы хрусталика приводят к его округлению, т.е. к увеличению его преломляющей силы (ссылки). Сторонники второй группы теорий считают, что изменение формы хрусталика для четкого видения близко расположенных объектов вызывается сместившимся вперед стекловидным телом, гидравликой внутриглазной жидкости задней камеры, натянутыми экваториальными связками хрусталика или сочетанием этих факторов.
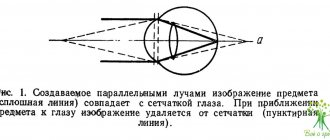
Серьезным аргументом в пользу сторонников интракапсулярного аккомодационного механизма стали результаты тщательно выполненной экспериментальной работы известного специалиста в области аккомодации глаза A. Glasser (1998, 1999). Он провел эксперимент на обезьянах с использованием ультразвуковой биомикроскопии (УБМ), чтобы определить, как именно изменяется экваториальный диаметр хрусталика на высоте напряжения аккомодации, т.е. в области ближайшей точки ясного зрения: увеличивается или уменьшается (рис.1). Если он увеличивается или не изменяется, то правы сторонники деформации экваториальной зоны хрусталика любым из описанных силовых воздействий – стекловидного тела, связочного аппарата или внутриглазной жидкостью. Если диаметр хрусталика уменьшается, то правы сторонники механизма изменения формы хрусталика за счет упругих сил (эластичности) его капсулы, т.е. внутрихрусталиковой структуры.
Эксперимент показал достоверное уменьшение экваториального диаметра хрусталика на высоте аккомодации, тем самым подтвердив интракапсулярную гипотезу Гельмгольца и его сторонников, придающих первостепенное значение в механизме аккомодации упругим силам хрусталиковых структур.
В то же время кроме доминирующей концепции «хрусталиковой» аккомодации существуют множество мнений об участии в этом процессе других структур глаза.
Чтобы полнее разобраться в механизме аккомодации, установить всех ее участников, разработать на этой основе эффективные средства клинической диагностики нарушений аккомодации и определить пути адекватной коррекции, в настоящее время используются три основных возможности: 1)компьютерное моделирование, 2)клинические наблюдения структур глаза с помощью современных средств визуализации и 3)определение тех или иных клинических показателей аккомодационной способности глаза.
Действительно, современные технические возможности позволяют создать весьма продуктивные виртуальные механико-математические (численные и аналитические) модели, которые становятся эффективным инструментом проверки научных гипотез и решения спорных вопросов, касающихся, в том числе, аккомодации.
В частности, использование биомеханической модели глаза (Иомдина Е.Н.) для изучения механизма аккомодации и его возрастных изменений показало, что с возрастом, при увеличении жесткости структур хрусталика и изменении соотношения жесткости ядра и коры хрусталика (при жесткости ядра выше жесткости коры) хрусталик теряет свою нормальную форму и деформационную способность, вследствие чего происходит не только возрастное падение объема аккомодации, но и изменение самого аккомодационного механизма: преломляющая сила глаза при аккомодации вблизь может оказаться меньше, чем при аккомодации вдаль, что не соответствует теории аккомодации Гельмгольца. В клинической практике эти возрастные особенности необходимо учитывать при подборе оптической коррекции, в частности, пресбиопической, планировании экстракции катаракты и определении параметров интраокулярной линзы (ИОЛ). После удаления содержимого хрусталика, включая жесткое ядро, аккомодационная способность глаза может возрасти. Подтверждением этого вывода, полученного на основе биомеханического моделирования, являются, в частности, недавно полученные результаты клинического исследования Овечкина И.Г. и соавт. (Овечкин И.Г., Беликова Е.И., Шалыгина Е.Л., Антонюк С.В., Овечкин Н.И. «Аккомодационная способность глаза у пациентов после факоэмульсификации катаракты с имплантацией монофокальных, мультифокальных и аккомодирующих интраокулярных линз». РОЖ 2014), которые показали появление истинной аккомодации (объективного аккомодационного ответа) у пациентов с пресбиопией после факоэмульсификации катаракты и имплантации монофокальных, и в особенности аккомодирующих ИОЛ. Этот вывод подтверждается динамикой базовых параметров аккомодационного ответа у оперированных пациентов и пациентов сопоставимой контрольной группы без патологии органа зрения. Полученные данные указывают на то, что аккомодирующие ИОЛ и (в значительно меньшей степени) монофокальные ИОЛ после удаления содержимого хрусталика и жесткого ядра пропорционально реагируют на сокращение цилиарной мышцы смещением вдоль оптической оси глаза.
Расчеты также показывают, что при максимальном напряжении аккомодации (т.е. при фокусировке на максимально близком и при этом четко видимом объекте) и при достаточно жестком хрусталике может наблюдаться провисание некоторых порций зонулярных волокон, что согласуется с результатами УБМ и может быть фактором риска развития подвывиха хрусталика.
Как было указано выше, большие возможности клинического наблюдения биомеханизма аккомодации предоставляют современные технологии визуализации структур глаза, такие как УБМ, оптическая когерентная томография (ОКТ), магнито- резонансная томография (МРТ) и Scheimpflug imaging».
На фотографиях, полученных методом «Scheimpflug imaging», ясно видно изменение радиуса кривизны хрусталика при аккомодации (рис.2 а, б), а также увеличение кривизны его передней поверхности в центральной зоне при аккомодации вдаль, вызванное наличием жесткого хрусталикового ядра (рис.2 в).
В.В. Страхов, используя УБМ (“Humphrey UBM- 840” с частотой колебания датчика 50 МГц), смогли выявить существенные возрастные топографо-анатомические изменения орбикулярного отдела задней камеры глаза, тонуса и направления хода цинновых связок, вызванные инволюционным увеличением размера хрусталика, что указывает, по мнению авторов, на важное значение уменьшения рабочего расстояния аккомодации в развитии пресбиопии (рис.3.). Автор делает вывод, что в биомеханизме пресбиопии, наряду с известными хрусталиковой (теория Гельмгольца) и мышечной (теория Дондерса) концепциями, появляется новая – связочная. Надо отметить, однако, что отсутствие трехмерной картины переднего отрезка глаза, которую невозможно получить с помощью двумерного УБМ сканирования, пока не позволяет поставить точку в изучении реального хода зонулярных волокон и объема передней и задней камеры глаза при аккомодации и требует дальнейших исследований этого вопроса.
Необходимо подчеркнуть, что вопрос, касающийся биомеханической роли связочного аппарата хрусталика в работе механизма аккомодации, является очень важным.
Как известно, зонулярные волокна цинновой связки подразделяются на три основные порции: передняя порция волокон (ППВ), задняя порция волокон (ЗПВ) и цилиоэкваториальные волокна (Рис.4.). Первые две порции волокон берут начало у зубчатой линии. Детальный вопрос о заднем креплении этих волокон обсуждается современными исследователями. ППВ прилежит к цилиарному телу, соединяясь с ним с помощью коротких тонких волокон вплоть до цилиарных отростков и, проходя группами между ними, прикрепляется к передней капсуле хрусталика в 1,5-2,0 мм от его экватора в отдельных фиксированных местах по окружности не равномерно, а сегментами.
Сегментарное по окружности прикрепление «пучков» ППВ ресничного пояска к капсуле хрусталика в принципе может позволить цилиарной мышце по-разному натягивать сегменты передней поверхности капсулы хрусталика, частично компенсируя естественный или приобретенный астигматизм оптической системы глаза. Возможно, при этом сегменты ЦМ могут сокращаться с различной интенсивностью. Эта гипотеза (о неравномерной аккомодации), сформулированная Светловой О.В. с соавт., ждет экспериментального подтверждения. ППВ толще и мощнее других порций зонулярных волокон. ЗПВ прилежит к передней пограничной мембране ЦТ, как бы дублируя ее своей сетью и соединяясь с ней системой переплетенных коротких тонких волокон до места прикрепления к задней капсуле хрусталика в 1,0 –1,5 мм от его экватора. Здесь ЗПВ вплетается в гиалоидно-капсулярную связку Вигера. Волокна ЗПВ тоньше, но многочисленнее, чем ППВ. Третья порция волокон, ЦЭВ, идущая от боковой поверхности оснований цилиарных отростков к экваториальной капсуле хрусталика, где они арочно и равномерно располагаются по экватору хрусталика вдоль линии их прикрепления. ЦЭВ слабее остальных зонулярных волокон и менее всего развиты. В зрелом возрасте они часто отмирают.
Участие связочного аппарата в уменьшении с возрастом аккомодационной способности подтверждается данными R.Michael et al. (2012), установившими снижение модуля упругости зонулярных волокон у людей старших возрастных групп.
Роли зонулярных волокон в нормальном аккомодационном процессе и развитии его нарушений посвящено исследование D.Goldberg (2011), который сформулировал теорию согласованного синхронного (взаимовозвратного) действия зонулярных волокон. Суть этой теории состоит в том, что при сокращении цилиарной мышцы передняя порция волокон ослабляется, а задняя натягивается и оказывает силовое воздействие на заднюю капсулу хрусталика, изменяя его толщину и форму задней поверхности. Во время расслабления цилиарной мышцы задние волокна ослабляются, и хрусталик сдвигается кзади из-за нарастающего напряжения передних связок (рис.5).
Кроме хрусталика и его связочного аппарата (зонулярных волокон), важнейшую роль в процессе аккомодации играет, безусловно, цилиарная мышца.
Цилиарная мышца, относящаяся к категории гладких мышц, условно разделяется на три порции (мышцы Брюкке, Иванова и Мюллера), в которых мышечные волокна ориентированы в разных направлениях (рис.6).
Меридиональная мышца Брюкке, самая мощная и длинная (в среднем 7 мм), прикрепляется к склере в области корнео-склеральной трабекулы и склеральной шпоры, свободно идет до зубчатой линии, где вплетается в хориоидею, доходя отдельными волокнами до экватора глаза. Общепринятым является мнение о парасимпатической иннервации этой мышцы («Аккомодация. Руководство для врачей п/ред. Катаргиной Л.А., Москва, 2012).
Порция внутренних волокон (циркулярная мышца Мюллера) не имеет прикрепления, наподобие сфинктера радужки, и располагается в виде кольца в самой вершине короны цилиарного тела. При ее сокращении вершина короны «заостряется», и отростки цилиарного тела приближаются к экватору хрусталика. Принято считать, что иннервация циркулярной мышцы так же, как и меридиональной, парасимпатическая.
Между этими двумя порциями располагаются радиальные мышечные пучки (радиальная мышца Иванова), которые часто образуют переплетающуюся решетку (Standring S., Borley N., Collins P., et al. «Gray’s Anatomy: The Anatomical Basis of Clinical Practice». Chapter 40. The eye. Elsevier. 2009). Эта порция составляет основную мышечную массу короны цилиарного тела и, имея прикрепление к увеальной части трабекулярной сети в прикорневой зоне радужки, свободно оканчивается в виде расходящегося радиально венчика на тыльной стороне короны, обращенной к стекловидному телу.
Очевидно, что при своем сокращении радиальные мышечные волокна, подтягиваясь к месту прикрепления, будут менять конфигурацию короны и смещать ее в направлении корня радужки. Несмотря на запутанность вопроса об иннервации радиальной мышцы, большинство авторов считают ее симпатической. Л.А.Деев с соавт. (1996) обнаружили не только наличие симпатических синапсов в цилиарном теле, но даже скопление адренэргических нервных терминалей в области радиальной порции цилиарной мышцы.
Биомеханизм, обеспечивающий весь объем аккомодации от дальнейшей до ближайшей точки ясного зрения и, наоборот, от ближайшей точки ясного зрения до дальнейшей находится под влиянием симпатического и парасимпатического отделов вегетативной нервной системы и реализуется через активный и пассивный мышечные компоненты за счет сокращения радиальной мышцы Иванова (симпатическая иннервация) и расслабления меридиональной мышцы Брюкке (парасимпатическая иннервация) и циркулярной мышцы Мюллера (парасимпатическая иннервация).
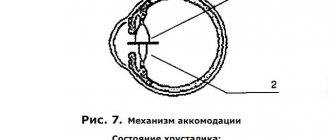
Вопрос о типе иннервации различных порций цилиарной мышцы важен для правильного биомеханического представления ее работы в процессе аккомодации, для понимания вклада каждой порции в аккомодационный механизм и для разработки средств целенаправленного регулирования их функциональной активности (рис.7).
В этом отношении плодотворным является применение медикаментозных моделей различных тонических состояний цилиарной мышцы. Так, В.В Страхов и соавт., используя инстилляции лекарственных средств (холинолитиков, адреномиметиков), активизирующих парасимпатические или симпатические рецепторы и тем самым воздействующих на различные порции цилиарной мышцы, с помощью УБМ зафиксировали изменение цилиарного тела и связочного аппарата хрусталика, подтвердив наличие активного механизма аккомодации глаза как для близи, так и для дали.
Несомненно, УБМ и другие технологии визуализации (УБМ, МРТ и др.) позволяют фиксировать изменение формы и геометрических параметров мышцы, ее смещение в процессе аккомодации, возрастные и другие особенности. Однако функциональные характеристики цилиарной мышцы, ее работоспособность, реакции на лекарственное или иное воздействие целесообразно также оценивать с помощью технических средств регистрации аккомодационного ответа.
Исследования показывают, что тонус цилиарной мышцы постоянно изменяется. Эти колебания называют аккомодационными микрофлюктуациями (АМФ). АМФ имеют определенную частоту и состоят из низко- и высокочастотного компонентов. Низкочастотный компонент (частота менее 0,6 Гц) является фоновым и не имеет клинического значения, а высокочастотный компонент (частота между 1,0 и 2,3 Гц) отражает флуктуации волокон цилиарной мышцы, и его измерение дает возможность оценить сократительную способность мышцы.
Для исследования АМФ используется аккомодограф Righton Speedy-K ver.MF-1, осуществляющий частотный анализ АМФ методом трансформации Фурье. Компьютерная аккомодография позволяет детально оценивать работоспособность цилиарной мышцы, диагностировать функциональное состояние аккомодации, ее динамические изменения, в том числе на фоне лечебных мероприятий.
Автоматическим и объективным является также исследование аккомодации с помощью бинокулярных авторефкератометров «открытого поля», например, Grand Seiko WR-5100K, WR-5500. Отличительной особенностью этих приборов является то, что благодаря открытому для взора испытуемого полю объект фиксации предъявляется в реальном (а не виртуальном) пространстве, предусмотрена возможность его перемещения от 20 см до бесконечности (5-6 м), а также приставления к глазам испытуемого линз различной диоптрийности, то есть измерения аккомодационного ответа в условиях дефокусировки. С помощью этой методики можно определить, в частности, тонус аккомодации, являющийся одним из важных показателей динамической рефракции. В темноте, т.е. при отсутствии стимула к аккомодации, некоторый тонус цилиарной мышцы сохраняется, за счет чего оптическая установка глаза соответствует точке, занимающей промежуточное положение между ближайшей и дальнейшей точкой ясного зрения. На положение этой точки, называемой также точкой покоя аккомодации и определяемой по разнице величин статической (т.е. в условиях циклоплегии) рефракции глаза и темнового фокуса аккомодации (т.е. рефракции в отсутствие аккомодационного стимула), могут влиять состояние вегетативной нервной системы (баланс симпатической и парасимпатической иннервации) и целый ряд других факторов. Определение этого показателя имеет большое значение как для выявления причин аккомодационных нарушений и связанных с этими нарушениями заболеваний глаз, в первую очередь, миопии, так и для определения адекватных средств их коррекции.
Применение комплекса методических приемов, включающих средства визуализации (УБМ), регистрацию аккомодационных показателей с помощью аккомодографии и бинокулярного авторефкератометра «открытого поля», позволило детально изучить влияние курса инстилляций адреномиметика — 2,5% раствора ирифрина (Сентисс, Индия) на состояние аккомодации в норме и при миопии. Ирифрин, содержащий в 1 мл 25 мг фенилэфрина гидрохлорида и 0,1 мг бензалкония хлорида, обладает выраженной альфа-адренергической активностью и при местном применении в офтальмологии суживает сосуды, расширяет зрачок, не вызывая циклоплегии. Научно- практический интерес к препарату обусловлен тем, что он является практически единственным существующим в настоящее время средством, стимулирующим аккомодацию для дали путем воздействия на симпатическую нервную систему.
Показано, что курсовое применение 2,5% ирифрина изменяет тонус цилиарной мышцы и баланс вегетативной иннервации в сторону снижения тонуса парасимпатической нервной системы, вызывает сдвиг динамической рефракции в сторону ослабления, способствует повышению запасов относительной аккомодации и объективного аккомодационного ответа, что можно объяснить действием отрицательной аккомодации (аккомодации для дали) и улучшением работоспособности цилиарной мышцы (Тарутта Е.П., Иомдина Е.Н., Тарасова Н.А., Филинова О.Б. «Влияние 2,5% ирифрина на показатели аккомодации и динамику рефракции у пациентов с прогрессирующей миопией». РОЖ, 2010, №2, с.30-33). При сочетании воздействия на симпатическую нервную систему (медикаментозная стимуляция ирифрином аккомодации для дали) с одновременным воздействием на парасимпатическую нервную систему (функциональные тренировки аккомодации при предъявлении стимула для близи) и физиотерапевтическим лечением, улучшающим состояние гемодинамики глаза, аккомодационная способность, нарушенная при миопии улучшается наиболее значительно (Волкова Е.М., Страхов В.В. «Применение Ирифрина как стимулятора аккомодации для дали». Клиническая офтальмология, 2005, № 2, с.86-88).
Полученные результаты подтверждаются данными аккомодографии на аппарате Speedy-K ver. MF-1 (Егорова А.В., Мыкольникова Е.С. «Препарат Ирифрин 2,5% в терапии компьютерного зрительного синдрома». Клиническая офтальмология, 2009, №1, 30-32).
В настоящее время ирифрин выпускается в виде глазных капель без консерванта (бензалкония хлорида), что существенно расширяет возможности его безопасного и эффективного использования для коррекции аккомодационных нарушений и профилактики прогрессирования миопии.
Кроме основных (активных) участников механизма аккомодации – хрусталика и его мышечно-связочного аппарата, принимает также участие хориоидея.
Макроскопически границей хориоидеи и цилиарного тела служит зубчатая линия, ora serrata, представляющей собой границу оптической части сетчатки. Хориоидея представляет собой тонкую мягкую эластичную коричневую мембрану, находящуюся в состоянии умеренного напряжения. Толщина хориоидеи в районе заднего полюса глаза составляет 0,22 мм, она постепенно истончается по направлению к периферии до 0,10 – 0,15 мм. На своем протяжении хориоидея прилежит к склере, а между ними находится узкая капиллярная щель – супрахориоидальное пространство. Задние концы более глубоко расположенных меридиональных волокон ЦМ переходят в эластические фибриллы хориоидеи. При сокращении меридиональных волокон ЦМ эта система эластических волокон натягивается. Расположенные более поверхностно волокна ЦМ своими задними концами входят в состав эпихориоидеи, системы тонких соединительнотканных пластинок, расположенных под склерой. Через них эти мышечные волокна фиксируются непосредственно к внутренней поверхности склеры. Далее кзади с помощью подобных, но более коротких пластинок к склере фиксируется и сама хориоидея.
Участие хориоидеи в механизме аккомодации представлено в работе (Светлова О., Макаров Ф., Котляр К., Засеева М., Кошиц И. Морфологические и функциональные особенности конструкции ресничного пояска хрусталика как ключевого звена в механизме аккомодации глаза человека. Морфология; 2003:7-16.) (рис.8). Авторы предлагают свое описание аккомодации вдаль (дезаккомодации) и вблизь, отводя хориоидее, по меткому определению А.И.Горбаня, роль своеобразной биологической пружины. При взгляде вдаль меридиональные и циркулярные волокна цилиарной мышцы расслабляются, ее радиальные волокна сокращаются, «растаскивая» циркулярные волокна подобно дилятатору радужки. Эластичная хориоидея возвращается (сжимается) в свое исходное (нерастянутое) состояние, оттягивая место заднего крепления волокон цинновой связки кзади. Передняя порция волокон цинновой связки при этом натягивается и прижимает хрусталик к стекловидному телу. При этом сам хрусталик вдавливается в переднюю пограничную мембрану (ППМ) стекловидного тела, но натянутая (и на этой фазе растянутая по поверхности ППМ) задняя порция волокон цинновой связки препятствует этому вдавлению. Хрусталик таким образом уплощается усилием передней порции волокон и прижимается к упруго-напряженной поверхности задней порции волокон и ППМ. Такое состояние стабильно и хорошо демпфирует возможные колебания хрусталика.
При взгляде вблизь меридиональные и циркулярные волокна цилиарной мышцы сокращаются, а ее радиальные волокна расслабляются. Цилиарное тело смещается кнутри-кпереди, растягивая эластичную хориоидею и таким образом подтягивая место заднего крепления волокон цинновой связки вперед. Натяжение передней и задней порций волокон цинновой связки ослабляется, и хрусталик, благодаря эластичности своей капсулы, принимает форму, более близкую к шарообразной. Необходимо отметить, что под действием разнонаправленных упругих сил: капсулы хрусталика с одной стороны и хориоидеи – с другой, передняя и задняя порции волокон цинновой связки во всех фазах аккомодации находятся в постоянно натянутом состоянии динамического равновесия. Степень их натяжения при аккомодации вблизь минимальна, а при аккомодации вдаль – максимальна. Цилиоэкваториальные волокна не участвуют в процессе аккомодации и служат, видимо, для поддержания формы хрусталика. Дело в том, что при аккомодации вблизь вещество хрусталика, стремящееся к шарообразной форме, имеет наименьший объем. При растяжении хрусталика его капсула растягивается. Внутренний объем увеличивается, а внутрихрусталиковое содержимое несжимаемо. Цилиоэкваториальные волокна растягивают капсулу у экватора, предотвращая возможное ее «скукоживание» или смещение внутрихрусталиковых масс относительно оптической оси.
В описании механизма аккомодации упоминается и стекловидное тело. Его участие в обеспечении оптической настройки глаза для четкого видения разноудаленных объектов является предметом активных дискуссий. Основная биомеханическая роль стекловидного тела, заполняющего всю внутреннюю полость глаза позади хрусталика, состоит в обеспечении нормального прилегания внутренних оболочек глаза друг к другу и к склере. По нашему мнению, стекловидное тело непосредственно, хотя и пассивно, участвует в процессе аккомодации, демпфируя и поддавливая хрусталик и позволяя ему незначительно смещаться вдоль передне-задней оси глаза.
Анализ различных теорий аккомодации свидетельствует о том, что еще одним фактором, определяющим поведение глаза при аккомодации, может являться жесткость склеры и ВГД. Однако расчеты, произведенные с использованием биомеханической модели, показывают, что объем аккомодации практически не зависит от механических свойств склеры, если ее продольный и окружной модули упругости находятся в диапазоне от 5 МПа до 40 МПа. Хотя цилиарная мышца и оказывает воздействие на склеру, ее поверхность при сокращении цилиарной мышцы смещается незначительно (смещение не превышает 0.02 мм). Не установлено и влияния на объем аккомодации изменений ВГД, если его величина составляет от 10 до 30 мм рт. ст. Однако при нарушении вязко-упругих свойств склеры, (например, при миопии) даже небольшая ее деформация при относительно повышенном ВГД может стать частично необратимой и привести к постепенному изменению формы корнеосклеральной капсулы глаза и к прогрессированию миопического процесса.
Многие специалисты по аккомодации высказывали мнение о том, что определенную роль в механизме аккомодации могут играть экстраокулярные мышцы, хотя большинство все же рассматривают их в качестве дополнительных участников аккомодационного процесса. Крайнюю точку зрения высказывал в этой связи N.V. Bates, который считал, что изменение ПЗО, вызванное давлением косых мышц на склеру, является основным механизмом аккомодации, а хрусталик вообще не изменяет форму.
Для изучения влияния экстраокулярных мышц на аккомодацию выполнена серия расчетов в рамках биомеханической модели глаза, в которых одновременно задавалось сокращение цилиарной и разных групп экстраокулярных мышц.
Моделирование показало, что существенное воздействие на объем аккомодации может оказать только избыточный тонус или рассогласованное сокращение экстраокулярных мышц, неравномерно сжимающих глазное яблоко при его повороте.
Подобное (рассогласованное) воздействие мышц приводит к деформации глазного яблока и к незначительному увеличению длины его переднезадней оси (ПЗО). Это, по-видимому, объясняется несимметричным креплением глазодвигательных мышц к склере и неравномерным (по распределению физико-механических свойств) строением склеры, в частности, ее повышенной жесткостью в местах крепления сухожилий. Увеличение ПЗО тем больше, чем ниже модуль упругости склеры и чем значительней рассогласованность сокращений экстраокулярных мышц, но в расчетах не удалось получить удлинения ПЗО более 0.5 мм.
Это означает, в частности, что нарушение сбалансированной работы глазодвигательных мышц может приводить к клиническим последствиям, связанным не только с косоглазием, но и с нарушением аккомодационной способности, а также с прогрессированием миопии, поскольку механическая устойчивость миопической склеры ниже, чем в норме (при эмметропии).
Моделирование работы экстраокулярных мышц показало, однако, что при их сокращении происходит некоторое смещение центра хрусталика относительно центра роговицы. Последним обстоятельством может объясняться то, что во время саккадических движений глаз зрительное восприятие снижено.
Этот результат совпадает с наблюдениями Гельмгольца: согласно его теории, при рассматривании предметов, расположенных на близком расстоянии, в человеческом глазу наряду с сокращением цилиарной мышцы, сужением зрачка, уменьшением глубины передней камеры происходит некоторое смещение хрусталика кпереди и книзу.
Несмотря на практически отсутствующее в норме влияние на величину ПЗО, повороты глаза (т.е. сокращения глазодвигательных мышц) в определенной степени вызывают отклонение оптической оси при сочетании хрусталиковой аккомодации и поворота глаза или просто при повороте, что может приводить к возникновению астигматизма, как компенсаторной реакции на отклонение оптической оси глаза. При этом собственно объем аккомодации изменяется незначительно. Учитывая, что в норме деформации экстраокулярных мышц не превышают 15%, максимальное падение объема аккомодации (расчетное значение для 35 лет) может составить около 1 дптр. Кратковременное возникновение искажения изображения, связанного с работой экстраокулярных мышц, скорей всего, остается незамеченным. В то же время продолжительные периоды искажений могут привести к необходимости их компенсации, что в свою очередь может вызвать развитие стойкого астигматизма оптической системы глаза.
Роль еще одной структуры глаза — роговой оболочки — в механизме аккомодации глаза представляется, согласно данным литературы, спорной и не вполне однозначной. Согласно гипотезе Гельмгольца, в норме роговица не изменяет свою оптическую силу и не участвует в акте аккомодации.